Caligus Species | Locality | Reference |
Caligus apodus (Brian, 1924) | Italy | Brian (1935) |
Caligus pageti Russell, 1925 | Egypt | Russell (1925) |
Caligus minimus Otto, 1821 | British Waters | Kabata (2003) |
Caligus mugilis Brian, 1935 | Italy | Brian (1935) |
Caligus bonito Wilson C.B., 1905 | North America | Cited by Paperna & Overstreet (1981) |
Caligus curtus Müller O.F., 1785 | Beaufort, North Carolina, USA | |
Caligus engraulidis Barnard, 1948 | Lake St.Lucia, South Africa | |
Caligus epidemicus Hewitt, 1971 | Mitchell River, Victoria, Australia | Hewitt (1971) |
Caligus irritans Heller, 1865 | Biscayne Bay, Florida | Skinner (1975) |
Caligus mutabilis Wilson C.B., 1905 | Palmyra Atoll, Central Pacific | Soler-Jiménez et al. (2019) |
Caligus orientalis Gusev, 1951 | Lake Mokoto | Urawa & Kato (1991) |
Caligus platytarsis Bassett-Smith, 1898 | Young off Heron Island, Queensland | Kabata (1965) |
Caligus praetextus Bere, 1936 | North America | Bere (1936) |
Caligus punctatus Shiino, 1955 | North America | Cited by Paperna & Overstreet (1981) |
Caligus rotundigenitalis Yü, 1933 | Taiwan | Ho & Lin (2004) |
Caligus rufimaculatus Wilson C.B., 1905 | Biscayne Bay, Florida | Skinner (1975) |
Caligus schistonyx Wilson C.B., 1905 | North America | Cited by Paperna & Overstreet (1981) |
Caligus schlegeli Ho & Lin, 2003 | Dongshih Fishing Port in Chiayi County | Ho & Lin (2003) |
CALIGUS TURKIYENSIS N. SP. (COPEPODA: CALIGIDAE) ON THE FLATHEAD GREY MULLET, MUGIL CEPHALUS (LINNAEUS) (MUGILIFORMES: MUGILIDAE) FROM THE SEA OF MARMARA, TURKEY
INTRODUCTION
Members of the Caligidae family are ectoparasite copepods on fish. There are 269 valid species listed in the genus Caligus, belonging to the family Caligidae (Walter & Boxshall 2024). Species belonging to Caligus are known as sea lice. These parasites are of global importance, because sea lice cause high levels of mortality as well as catastrophic losses in salmonid aquaculture. As a result, the direct and indirect effects of fish lice are economic losses in aquaculture (Johnson et al. 2004).
The number of Caligus species in the Mediterranean increased to 32, after the record of Caligus tunisiensis Hamdi, Benmansour, Zouari-Tlig, Kamanli, Özak, Boxshall, 2021 on Serranus scriba (Linnaeus, 1758) from the Mediterranean Sea. So far, 20 species of Caligus have been reported from the fishes of the Aegean Sea and Mediterranean Sea of Turkey: Caligus adenensis Özak, Sakarya & Boxshall 2019, Caligus apodus (Brian 1924), Caligus bonito C. B. Wilson 1905, Caligus brevicaudatus A. Scott 1901, Caligus dakari van Beneden 1892, Caligus diaphanus von Nordmann 1832, Caligus lagocephali Pillai 1961, Caligus lichiae Brian 1906, Caligus ligusticus Brian 1906, Caligus macrurus Heller 1865, Caligus minimus Otto, 1821, Caligus mulli Rodrigues, Özak, Silva & Boxshall 2018, Caligus pageti Russel 1925, Caligus pelamydis Krøyer, 1863, Caligus quadratus Shiino, 1954, Caligus scribae Essafi, Cabral & Raibaut 1984, Caligus solea Demirkale, Özak, Yanar & Boxshall 2014, Caligus temnodontis Brian 1924 and Caligus vexator Heller,1865. Five species of Caligus are known from the Sea of Marmara: C. apodus, C. bonito, C. diaphanus, Caligus minimus and Caligus zei Norman & T. Scott 1906 (Özak 2020).
Eighteen species of Caligus from Mugil cephalus Linnaeus, 1758 are listed in the world (Table I, WoRMS 2024), but only four of them are known on M. cephalus from the Mediterranean Sea. These species are C. apodus, C. minimus, C. mugilis and C. pageti (see Raibaut et al. 1971, Raibaut & Ben Hassine 1977, Raibaut et al. 1998, Kabata 2003). This paper describes a new species, Caligus turkiyensis n. sp. from the body surface and fins of the flathead grey mullet, M. cephalus, collected in the Sea of Marmara of Turkey. Two other Caligus species were identified in this survey: Caligus apodus and Caligus pageti on the body surface and fins of Chelon auratus (Risso, 1810) and M. cephalus from the Sea of Marmara, Turkey.
MATERIALS AND METHODS
One hundred and thirty specimens of the flathead grey mullet, Mugil cephalus and 63 of the golden grey mullet, Chelon auratus were caught by rod and line from Bandırma Bay, Turkey (the Sea of Marmara) in 2022. The parasites collected were fixed in 70 % ethanol. Some of the specimens were later cleared in lactic acid for 2 h before dissection of the appendages. The samples were dissected using an Olympus SZH10 and a Wild M5 stereomicroscope. The appendages were mounted on slides in a glycerin-gelatine mounting medium following Pritchard & Kruse (1982). The drawings of appendages were made with the aid of a drawing tube (Olympus® BH-DA). Microscope images were drawn on tracing paper, and the drawings were scanned and digitized. Additionally, microphotographs were taken with the aid of a Canon camera (EOS 1100D) connected to an Olympus CH30 phase contrast microscope. The scientific names and synonyms for parasites and hosts were checked with WoRMS (2024), and Froese & Pauly (2024), respectively. The nomenclature of copepod body parts follows Boxshall (1990) and Huys & Boxshall (1991). Samples of C. turkiyensis n. sp. in this study were compared with samples of Caligus minimus previously collected from Köyceğiz Dalyan Lagoon (Figs. 2, 6). Type and voucher specimens are deposited in the Zoology Collection of the İstanbul University, Faculty of Science, İstanbul, Turkey.
RESULTS
In this study, three species of Caligus were identified: Caligus apodus, Caligus pageti and Caligus turkiyensis n. sp.
Phylum Arthropoda von Siebold, 1848
Subphylum Crustacea Brünnich, 1772
Class Copepoda Milne Edwards, 1840
Order Siphonostomatoida Burmeister, 1835
Family Caligidae Burmeister, 1835
Genus Caligus Müller O.F., 1785
(Brian, 1924) (Fig.1)
Host: Chelon auratus(Risso, 1810) (Mugilidae) (n = 63)
Locality: Bandırma Bay (Balıkesir, Sea of Marmara, Turkey; 40°21’12.8” N 27°58’04.7” E); depth range: 1-3 m
Locality properties: A region where flows the sewage waters of Bandırma City Site on host: body surface, pectoral, ventral and caudal fins
Prevalence of infection: 21 %, Mean intensity: 1.3 Material examined: 1 female and 1 male deposited in the Zoology Collection of the Istanbul University, Faculty of Science (IUSHM 20231007-07, IUSHM 20231007-08)
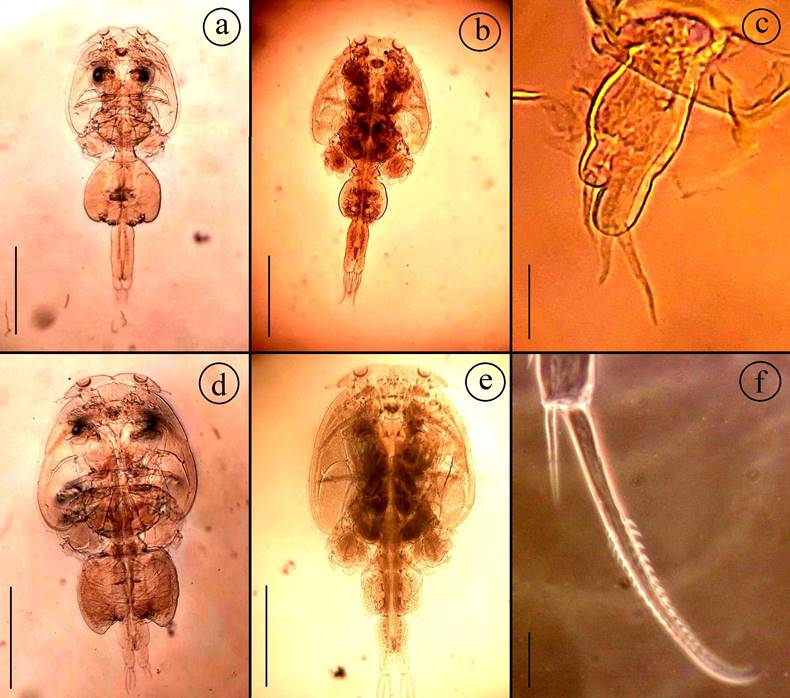
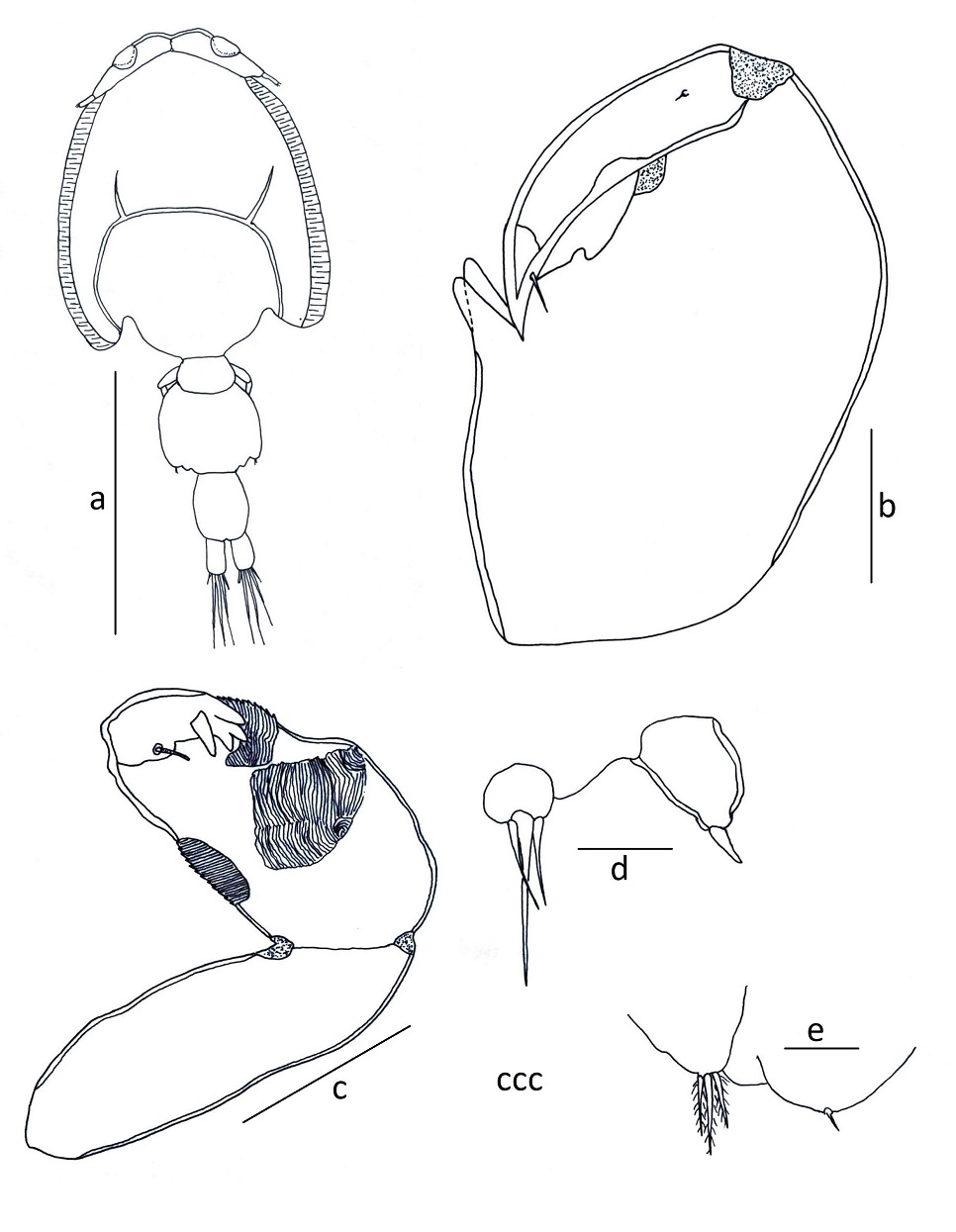
Adult female: Body (Fig. 1a) consists of a caligiform cephalothorax shield, a fourth pedigerous somite, a genital complex, and 1-segmented abdomen with paired caudal rami. Total body length 3.72 mm (3.33-4.26, n=4) caudal rami, excluding setae. Dorsal cephalothoracic shield is subcircular and slightly longer 1.74 mm (1.57-1.91) than wide 1.63 mm (1.40-1.81). Fourth pedigerous somite wider 0.37 mm (0.27-0.42) than long 0.21 mm (0.15-0.25), distinctly separated from the cephalothorax and genital complex. Genital complex subcircular, is wider 1.07 mm (0.84-1.23) than long 0.90 mm (0.73-1.12), about 1.3 times longer than the abdomen. Abdomen subrectangular and elongated, 1-segmented, longer 0.69 mm (0.66-0.72) than wide 0.37 mm (0.290.42). Caudal ramus subrectangular, slightly longer 0.19 mm (0.14-0.31) than wide 0.15 mm (0.11-0.20), each armed with six plumose setae. Egg sac long 2.39mm (1.98-3.21), wide 0.33 mm (0.31-0.42). Setal and spinal formula of 1-4 legs are given in Table II.
Adult male: Body (Fig. 1b) consists of a caligiform cephalothorax shield, fourth pedigerous somite, genital complex and 1-segmented abdomen with paired caudal rami. Total body length 3.57 mm, (3.52-3.62, n = 2) caudal rami, excluding setae. Dorsal cephalothoracic shield subcircular, slightly longer 1.91 mm (1.901.92) than wide 1.72 mm (1.70-1.74). Fourth pedigerous somite wider 0.42 mm (0.41-0.42) than long 0.22 mm (0.21-0.22), distinctly separated from the cephalothorax and genital complex. Genital complex subcircular, is slightly wider 0.71 mm (0.7-0.72) than long 0.61 mm (0.590.62), about as long as the abdomen. Abdomen subrectangular 1-segmented, longer 0.62 mm (0.60-0.63) than wide 0.32 mm (0.31-0.32). Caudal ramus subrectangular, slightly longer 0.23 mm (0.22-0.23) than wide 0.12 mm (0.110.12) and each armed with six plumose setae.
Remarks: Brian (1924) first described C. apodus (as syn Pseudocaligus apodus) on M. cephalus from Mauritania. This species is easily distinguished from the other caligid species by the absence or presence of the vestigial fourth leg in female and male samples. Ben Hassine (1983) mentioned the presence of vestigial fourth legs in both females and males of C. apodus from Tunisia. Özak et al. (2013) did not observe a vestigial fourth leg in female samples from Turkey. They also found a minute, rod-like trace of a possible fourth leg in one of the two females in Brian’s (1935) samples from Italy. No evidence of the fourth leg in female individuals was found in the present study.
Özak et al. (2013) also compared male samples of C. apodus from Turkey and Brian’s (1935) samples from Italy. Özak et al. (2013) found three unequal and spiniform elements at the apex in the samples from Turkish Mediterranean Sea Coasts and two spiniform terminal elements in Italian material. The fourth leg with three terminal spiniform elements in the male samples (Fig. 1c) in this study is also compatible with those of Özak et al. (2013).
The total body length of females in this study was smaller than in Brian’s (1935) length of 4.3-5.00 mm and Özak et al.’s (2013) of 4.30 mm. The total body length of males in this study was greater than Özak et al.’s (2013) of 3.19 mm and smaller than in Brian’s (1935) of 3.19-4.00 mm.
Russel, 1925 (Fig. 1)
Host: Mugil cephalus Linnaeus, 1758 (Mugilidae) (n = 130)
Locality: Bandırma Bay (Balıkesir, Sea of Marmara, Turkey; 40°21’12.8”N 27°58’04.7”E); depth range: 1-3 m
Locality properties: A region where flows the sewage waters of Bandırma City Site on host: body surface, pectoral, ventral and caudal fins
Prevalence of infection: 10 %, Mean intensity: 1.2 Material examined: 1 female and 1 male deposited in the Zoology Collection of the Istanbul University, Faculty of Science (IUSHM 2023100709, IUSHM 20231007-10)
Adult female: Body (Fig. 1d) consists of a caligiform cephalothorax shield, a fourth pedigerous somite, a genital complex and 1-segmented abdomen with paired caudal rami. Total body length 4.17 mm (3.42-5.07, n = 4). Dorsal cephalothoracic shield subcircular, is slightly longer 2.40 mm (2.02-2.93) than wide 2.28 mm (1.87-2.81). Fourth pedigerous somite wider 0.64 mm (0.51-0.81) than long 0.25 mm (0.22-0.29), distinctly separated from the cephalothorax and genital complex. Genital complex subrectangular, wider 1.35 mm (0.911.71) than long 0.89 mm (0.62-1.12); about 2.3 times longer than the abdomen, and with slightly lobate posterolateral corners of genital complex and extending to middle of abdomen. Abdomen subrectangular, 1-segmented, slightly wider 0.44 mm (0.38-0.51) than long 0.39 mm (0.35-0.46). Caudal ramus subrectangular, longer 0.24 mm (0.21-0.27) than wide 0.16 mm (0.120.20), each armed with six plumose setae. Egg sac long 2.79 mm (1.72-3.62), wide 0.39 mm (0.31-0.42). Setal and spinal formula of 1-4 legs are given in Table II.
Adult male: Body (Fig. 1e) consists of a caligiform cephalothorax shield, a fourth pedigerous somite, a genital complex and 1-segmented abdomen with paired caudal rami. Total body length 4.49 mm (4.21-4.63, n = 3). Dorsal cephalothoracic shield subcircular, slightly longer 2.63 mm (2.41-2.75) than wide 2.57 mm (2.31-2.72). Fourth pedigerous somite, wider 0.66 mm (0.61-0.69) than long 0.32 mm (0.31-0.33), distinctly separated from the cephalothorax and genital complex. Genital complex subcircular, wider 0.98 mm (0.91-1.02) than long 0.70 mm (0.67-0.72), about 1.3 times longer than abdomen. Abdomen subrectangular and 1-segmented, slightly wider 0.55 mm (0.51-0.61) than long 0.52 mm (0.51-0.52). Caudal ramus subrectangular, longer 0.32 mm (0.31-0.34) than wide 0.22 mm (0.22-.0.24), each armed with six plumose setae.
Remarks:Caligus pageti was first described on M. cephalus from Egypt by Russel (1925). The most distinguishing characteristic of C. pageti is the fourth leg, which is serrated to middle of inner and outer margins of fourth leg. The fourth leg is 3-segmented, with a protopodal segment and 2-segmented exopodal segment; protopodal segment with a plumose first exopodal segment with outer spine; second exopodal segment with long outer apical spine and one subapical spine about one-third the length of the apical spine; long apical spine serrated to the middle of inner and outer margins (Fig. 1f).
n. sp. (Figs. 2-6)
Type host: Mugil cephalus Linnaeus, 1758(Mugilidae) (n = 130)
Locality: Bandırma Bay (Balıkesir, Sea of Marmara, Turkey; 40°21’12.8”N 27°58’04.7”E); depth range: 1-3 m
Locality properties: A region where flow the sewage waters of Bandırma City
Site on host: body surface, fins
Prevalence of infection: 22 %, Mean intensity: 1.1 Type Material: Holotype female and paratype male deposited in the Zoology Collection of the Istanbul University, Faculty of Science (IUSHM 20231007-03, IUSHM 20231007-04)
Etymology: The species name refers to Turkey, close to where the material was collected.
Description
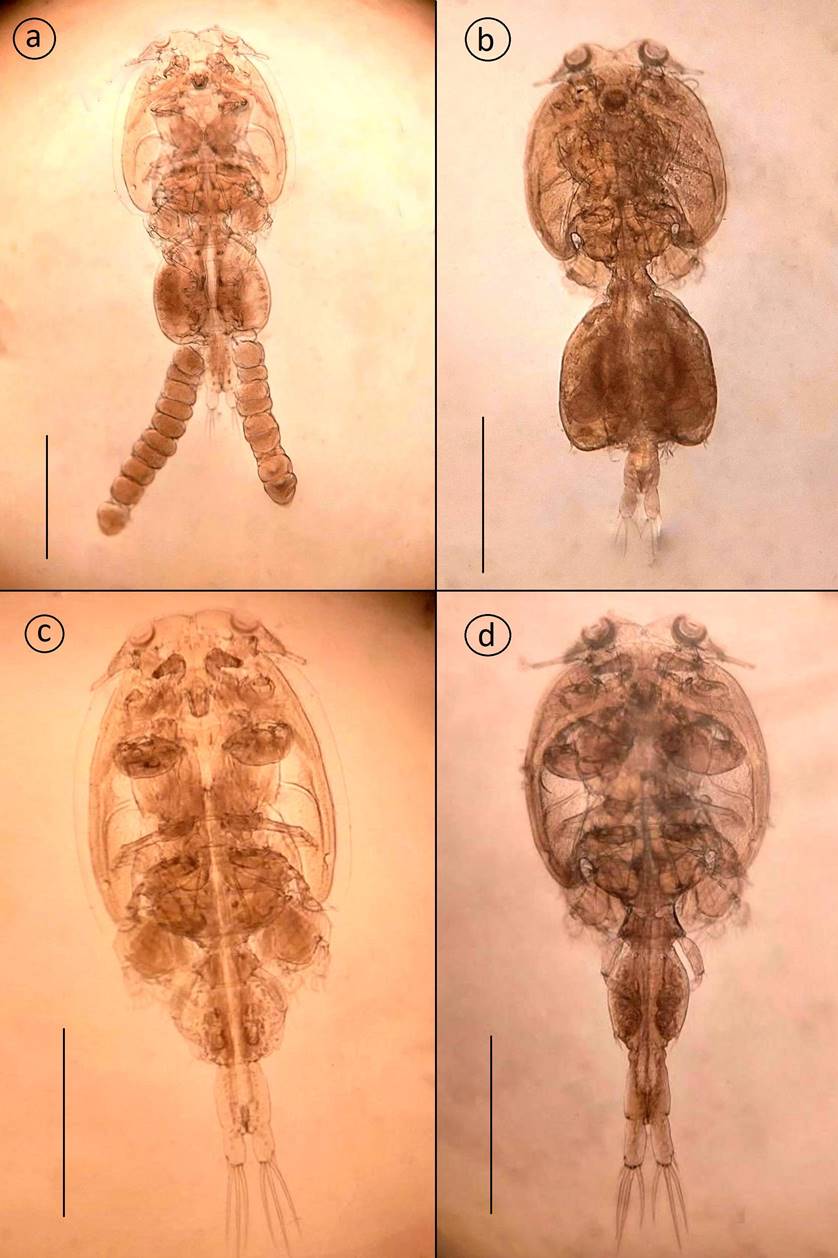
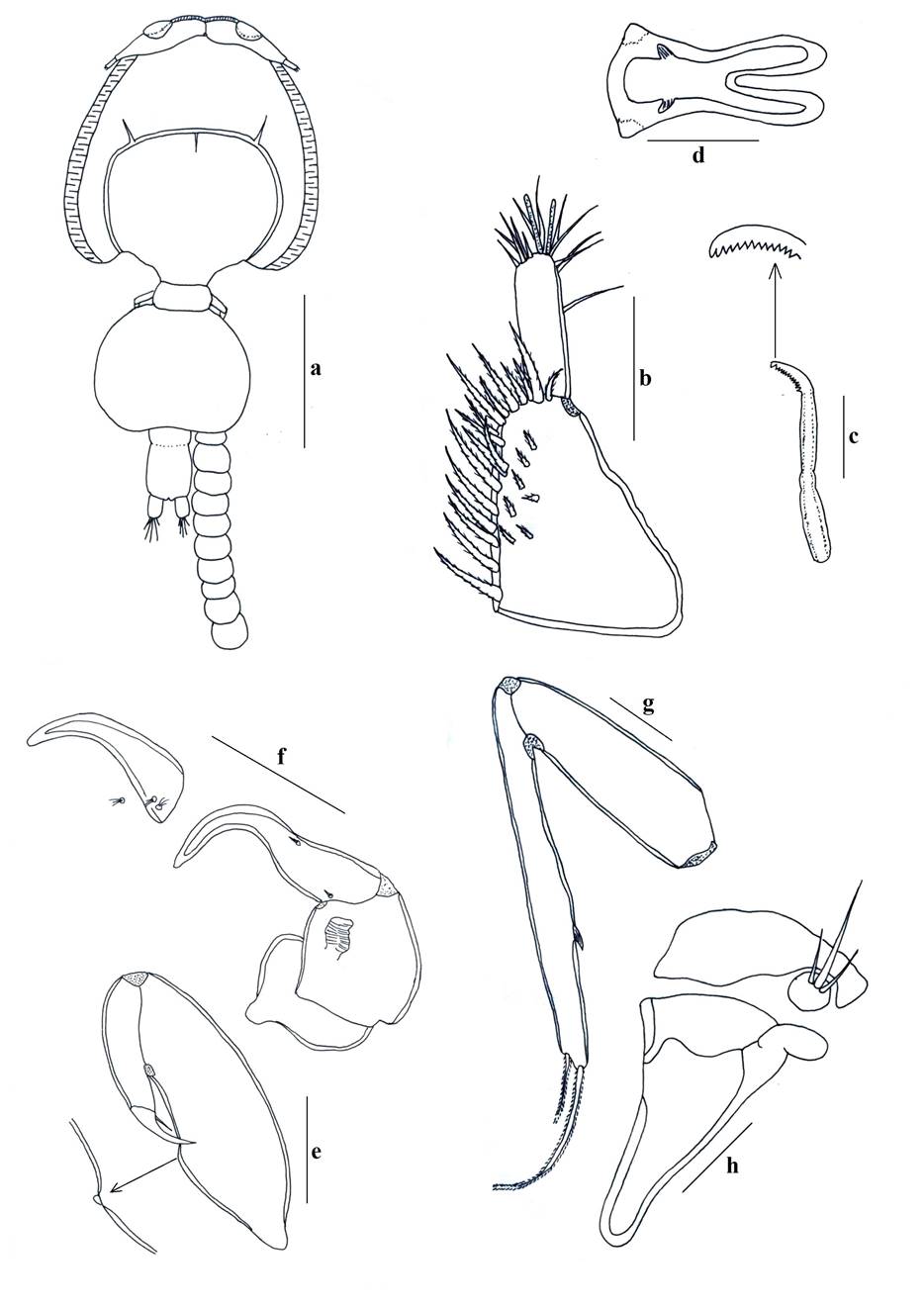
Adult female: Cephalothorax subcircular, dorsoventrally flattened, lateral zones of dorsal cephalothoracic shield associated with welldeveloped marginal membranes. Body consists of a caligiform cephalothorax shield, a fourth pedigerous somite, a genital complex and an indistinctly 2-segmented abdomen with paired caudal rami. Total body length 3.79 mm (3.134.61, n = 7) (Fig. 2a, 3a). Dorsal cephalothoracic shield subcircular, longer at 1.82 mm (1.52-2.01) than wide 1.61 mm (1.34-1.73). Fourth pedigerous somite is wider at 0.46 mm (0.24-0.61) than long 0.23 mm (0.14-0.32), distinctly separated from the cephalothorax and genital complex. Genital complex subcircular, is slightly wider at 1.04 mm (0.77-1.52) than long 0.94 mm (0.741.31), about 1.6 times longer than the abdomen and with nonlobate posterolateral corners of genital complex. Abdomen subrectangular, indistinctly with two somites, first abdominal somite slightly wider at 0.31 mm (0.21-0.41) than long 0.18 mm (0.12-0.22), anal somite longer at 0.41 mm (0.280.48) than wide 0.33 mm (0.28-0.42). Egg sac 2.11 mm (1.82-2.51) long by 0.36 mm (0.31-0.42) wide.
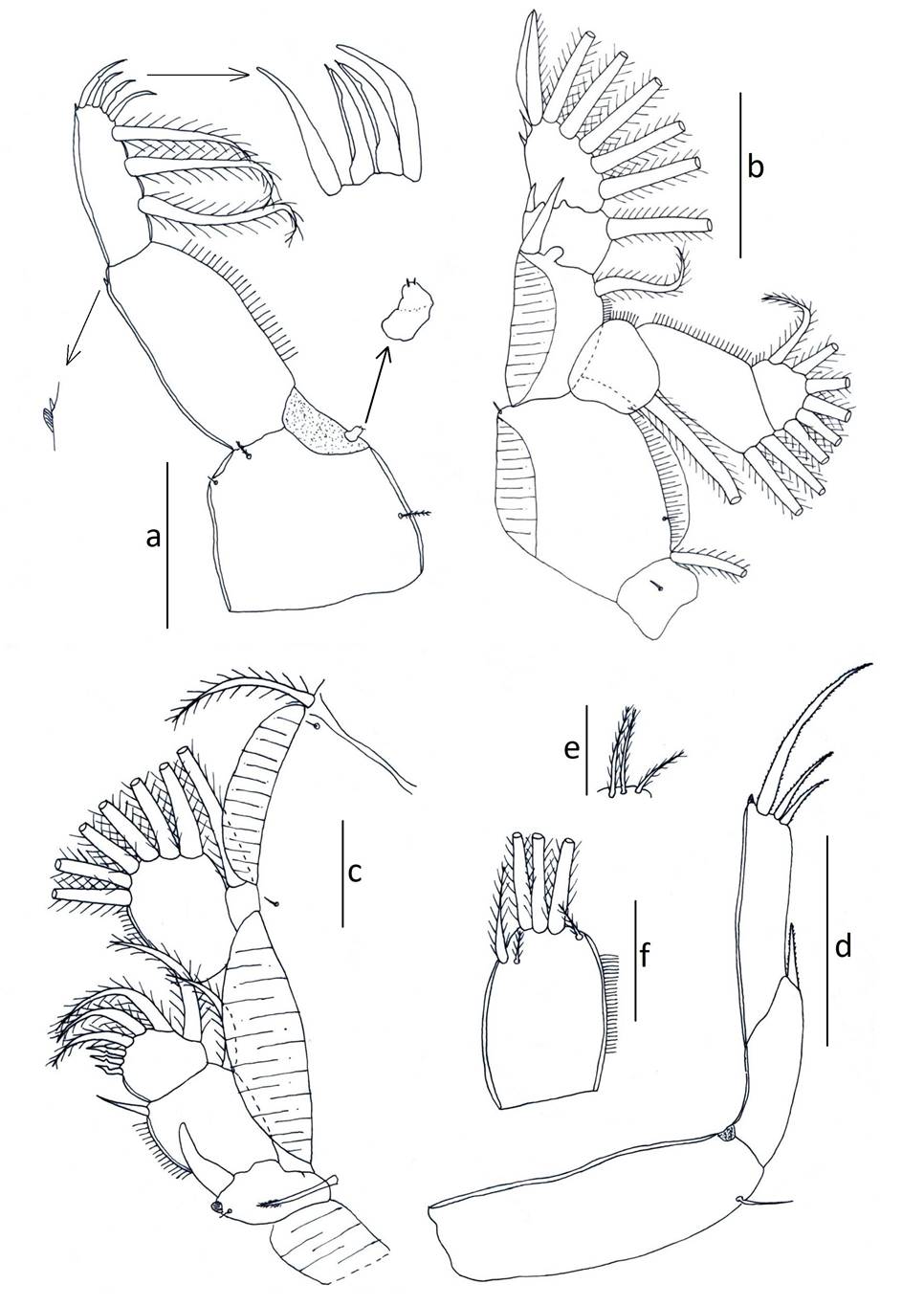
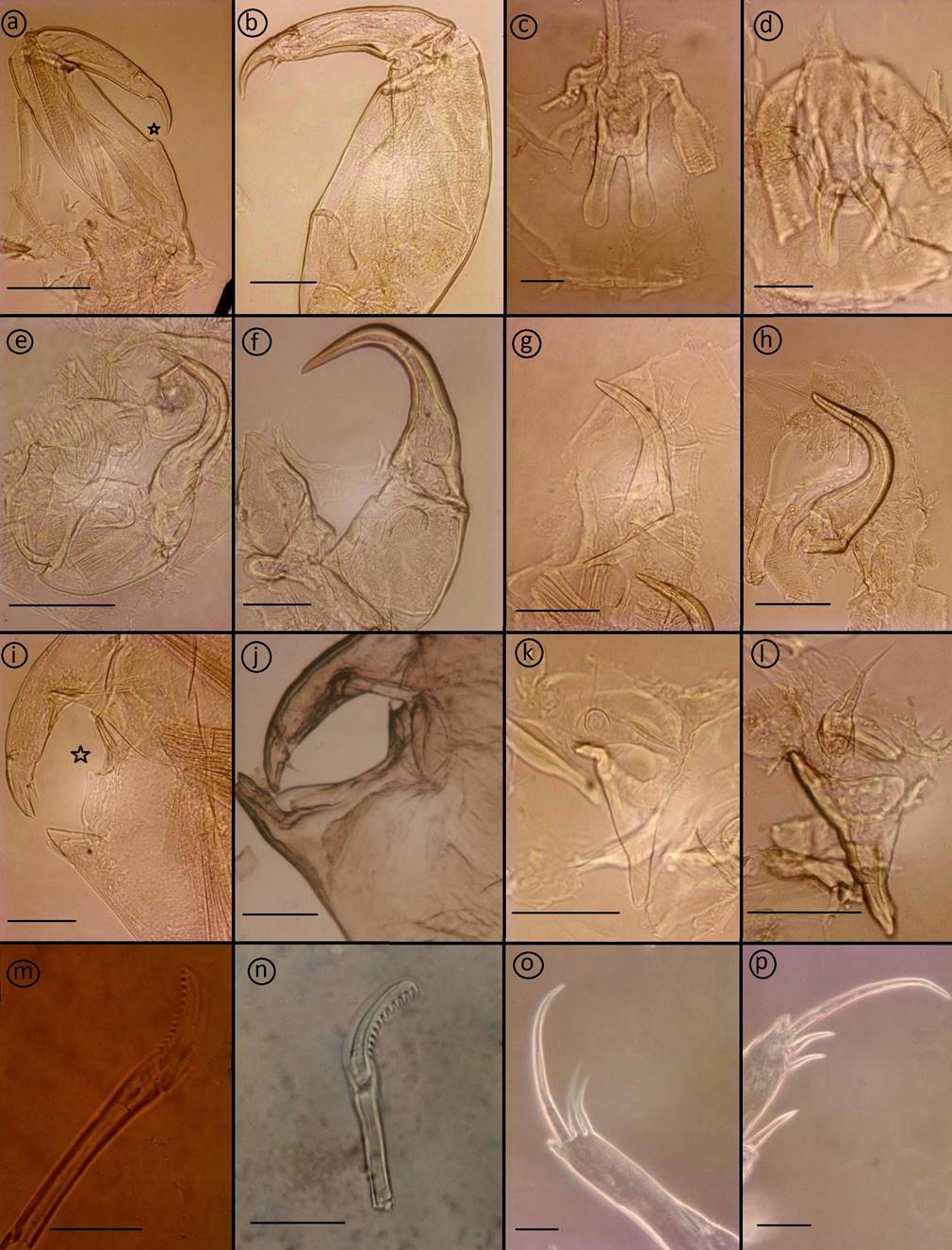
Antennule (Fig. 3b) 2-segmented; proximal segment distinctly larger than distal, armed with 24 setae; distal segment armed with isolated subterminal seta on posterior margin and 11 naked setae plus two aesthetascs around the apex. Antenna (Fig. 3f, 6e) uniramous, 3-segmented; proximal segment subrectangular with rounded margins and a short process; middle segment subrectangular and armed with a large corrugated adhesion pad on the dorsal surface; terminal segment forming a recurved claw armed with a seta near the middle area and posterior margin. Post-antennal process (Fig. 3f, 6g) well-developed, slender and slightly curved, ornamented with two trisensillate papillae on the basal part and a single similar trisensillate papilla on the adjacent ventral cephalothoracic surface near the base of the post-antennal process. Mandible (Fig. 3c, 6m) stylet-like, armed with 13 marginal teeth on inner side near the apex. Maxillule (Fig. 3h, 6k) comprising an anterior papilla bearing three naked setae (one medial long seta and two lateral small setae), and an anterior process with folding, and posterior process slender with rounded margin. Maxilla (Fig. 3g) 2-segmented and brachiform; proximal segment (lacertus) unarmed; distal segment (brachium) slender, bearing subterminal hyaline membrane on the inner margin plus short canna and long calamus distally. Maxilliped (Fig. 3e, 6a) subchelate, protopodal segment (corpus) elongate and bearing a slight expansion on myxal surface; distal subchela representing endopodal segments plus claw; small seta at the concave margin of claw base. Sternal furca (Fig. 3d, 6c) with subrectangular box, elongated and parallel tines with little divergence. Leg 1 (Fig. 4a) biramous with sympods joined by a slender intercoxal sclerite; sympod with inner and outer plumose seta, plus a setule; and endopod represented by a process on the posterior margin of basis, bearing two-minute vestigial setae on the apex. Exopod 2-segmented, with first exopodal segment robust, about 2.7 times longer than wide, armed with a small outer spine at the distal corner and ornamented with a row of setules along the mid-section of the posterior margin. Distal exopodal segment with three plumose setae posteriorly plus four distal margin elements; spine 1 (outermost) and seta 4 smooth and longer than spines 2-3; spines 2 and 3 about equal in length, both lacking accessory process and arcuated with blunt ends. Leg 2 (Fig. 4b) biramous, with distinct coxa and basis; coxa short, bearing a long plumose inner seta and a sensillium on the ventral surface; basis armed with a short naked seta on the outer distal corner plus extensive marginal membrane along the posterior margin and a sensillium near the mid-length of the anterior margin. Exopod 3-segmented: first exopodal segment with inner plumose seta, an outer spine, extending to the middle of the third exopodal segment and with a row of setules on the inner margin; the second exopodal segment is the smallest, with an inner plumose seta and an outer distal spine and not extending across to the anterior margin of the third segment. Third exopodal segment with three outer spines and five inner plumose setae; first spine simple and smallest; second spine slightly longer than first spine and third spine with an outer marginal membrane and fine setules along the inner margin. Endopod 3-segmented; first endopodal segment subrectangular, armed with an inner plumose seta and ornamented with a row of setules along the outer margin; second segment elongated, armed with two inner plumose setae plus ornamented with a row of setules along the outer margin; third segment smallest, with six plumose setae. Leg 3 (Fig. 4c) protopodal part flattened and joined by plate-like intercoxal sclerite, forming an apron ornamented with marginal membrane posteriorly and along the lateral margin anterior to the exopod: bearing inner plumose seta at the junction with intercoxal plate and an outer plumose seta dorsal to the base of the exopod; sensillae located adjacent to the inner coxal seta and adjacent to the origin of endopod. Exopod 3-segmented: first segment with a large, recurved outer spine; a hyaline flap and sensillium along the corner of spine; second segment with an outer spine, inner plumose seta and setules along outer margin; the third segment with three outer spines (third spine slightly longer than the other two) and four short plumose setae. Endopod 2-segmented: first segment is armed with a long inner plumose seta; second segment is armed with six plumose setae, ornamented with rows of setules along the outer margin. Leg 4 (Fig. 4d, 6o) uniramous. Protopodal segment with an outer seta derived from the basis. Exopod 2-segmented; first segment bearing a short, slender, outer distal spine, not extending to the middle part of second exopodal segment; second segment with three apical spines increasing in length from outer to inner; first spine slightly shorter than second and third spine three times as long as. Leg 5 (Fig. 4e) is represented by three plumose setae. Setal and spinal formula of 1-4 legs are given in Table II. Caudal ramus (Fig. 4f) subrectangular, longer 0.19 mm (0.13-0.27) than wide 0.14 mm (0.10-0.25), each armed with three distal setae the longest on the outer margin, a median size seta, and other the two located opposed, and with a row of setules on inner margin.
Adult male: Cephalothorax subcircular, dorsoventrally flattened, lateral zones of dorsal cephalothoracic shield associated with welldeveloped marginal membranes. Body consists of a caligiform cephalothorax shield, a fourth pedigerous somite, a genital complex and 1-segmented abdomen with paired caudal rami. Total body length 3.67 mm (3.30-4.69, n = 7) (Fig. 2c, 5a). Dorsal cephalothoracic shield subcircular, is slightly longer 2.12 mm (1.82-2.71) than wide 1.75 mm (1.52-2.22). Fourth pedigerous somite at wider 0.49 mm (0.41-0.61) than long 0.29 mm (0.21-0.35), distinctly separated from the cephalothorax and genital complex. Genital complex subcircular, is slightly at wider 0.69 mm (0.56-0.92) than long 0.56 mm (0.46-0.72), and about 1.2 times longer than abdomen. Abdomen subrectangular, 1-segmented, longer at 0.46 mm (0.41-0.60) than wide 0.34 mm (0.30-0.42). Caudal ramus subrectangular, longer at 0.23 mm (0.200.31) than wide 0.13 mm (0.11-0.22), each armed with six plumose setae. Antennule, mandible, post-antennal process and maxilla as in female. Maxilliped (Fig. 5b, 6i) protopod (corpus) is slightly swollen and armed with one small process at distal part and one big process as bifid at posterior part of myxal surface opposing tip of subchela; distal subchela armed with a short seta at the concave margin of claw base and a spine at posterior margin. Antenna (Fig. 5c) 3-segmented; proximal segment elongated and unarmed; middle segment reflexed, swollen proximally, with corrugated adhesion pads on medial and distal surfaces; distal segment with four plates on trifid direction and with basal seta. Maxillule (Fig. 5d) comprising an anterior papilla bearing three naked setae (one long and two small) and reduced posterior process. Legs 1 to 4, as in female. Leg 5 and 6 (Fig. 5e) are represented by three plumose setae and a minute spine, respectively.
Remarks: The report of C. mugilis from M. cephalus caught in Bandırma Bay (Sea of Marmara) by Alaş & Öktener (2017) is erroneous according to Özak (2020) for the following reasons: the female had an abdomen as long as the genital complex; the female maxilliped had a distinct, subtriangular myxal process; the posterolateral corners of the genital complex were indistinctly lobate and did not extend beyond the posterior margin of the genital complex. This study provided an opportunity to collect and examine the specimens of Alaş & Öktener (2017) again. The morphology of the new species appears to have the closest similarity to the four species Caligus curtus Müller O.F., 1785, Caligus dicentrarchi Cabral & Raibaut, 1986, C. minimus and C. mugilis (Table IV). Caligus mugilis was described for the first time from the mucous membrane of the mouth of M. cephalus from Portoferraio, Italy by Brian (1935), but he considered it to be a subspecies of Caligus minimus and named it as Caligus minimus var. n. mugilis. Later, Raibaut & Ben Hassine (1977) accepted C. mugilis as a different species from Chelon labrosus (Risso, 1827) and described it from only female specimens. They also compared females of C. mugilis from C. labrosus and C. minimus from Dicentrarchus labrax (Linnaeus, 1758) in Tunisia. The female of the new species differs from C. mugilis in having a smaller body at 3.79 mm (5.33 mm in C. mugilis); a genital complex which is about 1.6 times longer than the abdomen (3.2 times in C. mugilis); an abdomen with two indistinct somites (one somite in C. mugilis); tines of sternal furca little divergent, slightly elongated (divergent, short tines in C. mugilis); number of teeth on mandible as 13 (12 in C. mugilis); posterolateral corners of genital complex indistinctly lobate not extending to middle or anterior of the abdomen (distinctly lobate, extending slightly beyond the middle of abdomen in C. mugilis); ratio of length/anterior margin of maxillule endopodite is 1.45 (1.1 in C. mugilis); myxal surface of maxilliped with a slight process (absent in C. mugilis); and ratio of first spine/third spine on fourth leg is 3.2 (3.4 in C. mugilis). The male of the new species could not be compared as the male of C. mugilis was not described. The other species reported from M. cephalus is Caligus minimus which was described on D. labrax by Otto (1821). The female of the new species differs from C. minimus in having a smaller body 3.79 mm in comparison with 4.38 mm in C. minimus; genital complex is about 1.6 times longer than abdomen (2.3 times in C. minimus); abdomen with indistinctly two somites (one somite in C. minimus); tines of sternal furca are little divergent, slightly elongated (slightly divergent short in C. minimus) (Fig. 6d); number of teeth on mandible is 13 (12 teeth in C. minimus) (Fig. 6m); posterolateral corners of genital complex indistinctly lobate not extending to middle or anterior of the abdomen (distinctly lobate, extend slightly beyond one third of abdomen in C. minimus); terminal segment of antenna and postantennal process slightly recurved (strongly recurved in C. minimus) (Fig. 6f, h); ratio of length/anterior margin of maxillule endopo.
Species | Leg 1 (exopod/ endopod) | Leg 2 (exopod/ endopod) | Leg 3(exopod/ endopod) | Leg 4 (exopod/ endopod) |
C. turkiyensis n. sp | I-0; III, 1, 3/ vesitigial | I-1; I-1; II, I, 5/ 1-0; 2-0; 6 | I,0; I-1; III, 4/ 1-0; 6 | I-0; III/ Absent |
C. apodus | I-0; III, I, 3/ vestigial | I-1; I-1; II, 1, 5/ 0-1; 0-2; 6 | I-0; I-1; III, 4/ 0-1; 6 | Absent |
C. pageti | I-0; III, 1, 3/ vestigial | I,1; I,1: II, I, 5/ 1,0; 2,0; 6 | I,0; I-1; III,4/ 1-0; 6-0 | I-0; II/ Absent |
Male of the new species differs from C. minimus in having a smaller body 3.67 mm (4-5 mm in C. minimus); myxal surface of maxilliped with one small process in distal and one large bifid process in proximal (one large process in proximal of C. minimus) (Fig. 6j); terminal segment of antenna with quadrifid plate and a basal seta (quadrifid plate in C. minimus).
Caligus curtus was described from Merlangius merlangus (Linnaeus, 1758) in Denmark and Norway waters by Müller (1785). It can be distinguished from C. turkiyensis n. sp. in having: a greater body, 8.7 mm in comparison to 3.79 mm; a genital complex about 2.5 times longer than the abdomen (1.6 times in C. turkiyensis n. sp.); an abdomen with one somite (indistinctly two somites in C. turkiyensis n. sp.); tines of sternal furca slightly divergent, short parallel (little divergent, slightly elongated in C. turkiyensis n. sp.); number of teeth on mandible 12 (13 teeth in C. turkiyensis n. sp.); posterolateral corners of genital complex distinctly lobate, extend slightly beyond one third of abdomen (indistinctly lobate not extending to middle or anterior of the abdomen in C. turkiyensis n. sp.); terminal segment of antenna and postantennal process strongly recurved (slightly recurved in C. turkiyensis n. sp.); ratio of length/anterior margin of maxillule endopodite of 0.8 (1.45 in C. turkiyensis n. sp.); and ratio of first spine/third spine on fourth leg of 5 (3.2 in C. turkiyensis n. sp.). Male of C. curtus differs from C. turkiyensis n. sp. in having a greater body of 7.93 mm in comparison to 3.67 mm in C. turkiyensis n. sp.; myxal surface of maxilliped with one large process in both proximal and distal (one small process in distal and one large bifid process in proximal of C. turkiyensis n. sp.); and terminal segment of antenna with bifid plate and a basal seta (quadrafid plate and a basal seta in C. turkiyensis n. sp.) and denticles on first spine of fourth leg present (absent in C. turkiyensis n. sp.). Caligus dicentrarchi was found on D. labrax farmed in Corsica and Sete by Cabral & Raibaut, 1986. The female of the new species can be differentiated from C. dicentrarchi in having: a smaller body of 3.79 mm in comparison to 6.2 mm in C. dicentrarchi; genital complex is about 1.6 times longer than the abdomen (2.6 times in C. dicentrarchi); abdomen with indistinctly two somites (one somite in C. dicentrarchi); tines of sternal furca little divergent, slightly elongated (short parallel in C. dicentrarchi); number of teeth on mandible 13 (12 teeth in C. dicentrarchi); posterolateral corners of genital complex indistinctly lobate not extending to middle or anterior of the abdomen (distinctly lobate, extend slightly beyond one-third of abdomen in C. dicentrarchi); ratio of length/anterior margin of maxillule endopodite of 1.45 (1.2 in C. dicentrarchi); myxal surface of maxilliped with a slight process (absent in C. dicentrarchi); and ratio of first spine/third spine on fourth leg of 3.2 (4.5 in C. dicentrarchi). Male of the new species differs from C. dicentrarchi in having a smaller body of 3.67 mm (6.5 mm in C. dicentrarchi); myxal surface of maxilliped with one small process in distal and one large bifid process in proximal (one large process in proximal of C. dicentrarchi); terminal segment of antenna with quadrifid plate and a basal seta (trifid plate in C. dicentrarchi). Boxshall (2018) classified some Caligus species in 5 groups according to a combination of several morphological characters, such as leg 1, 2 and 4, antenna, maxilla, and abdomen of female. These groups are Caligus bonito-group, Caligus confusus-group, Caligus diaphanus-group, Caligus macarovi-group, and Caligus productus-group. The number of these groups reached 7 with the revision of Caligus pseudorhombi-group and Caligus undulatus-group by Ohtsuka & Boxshall (2019) and Ohtsuka et al. (2020), respectively. Caligus turkiyensis n.sp. may also be closely related to the C. macarovi-group. First and second exopodal segments of 3-segmented leg 4 with I and III spines in this group are the same in the new species. The proximal segment of female antenna has a posterior process, as in this group. The new species differs from the C. macarovi-group: female abdomen is 2-segmented indistinctly in the new species, but group carries 1-segmented. Distal of maxilla brachium in this group has marginal serrations, but is absent in the new species. Structure of setal and spinal of leg 1 in new species is similar to the rest group. This species has also the distal exopodal segment of leg 1 bears 3 plumose setae and 4 distal elements. Spines 1-3 are all subequal in length and seta 4 is markedly longer than spines in this group. Spine 1 (outermost) and seta 4 are smooth and longer than spines 2-3; spines 2 and 3 are about equal in length in new species. Only spines 2 and 3 carry accessory processes in this group, but spines 2 and 3 both lack accessory process and arcuated with blunt ends. Many morphological characters of the new species are different from the C. macarovi-group. Therefore, it is likely that it does not belong to this group.
DISCUSSION
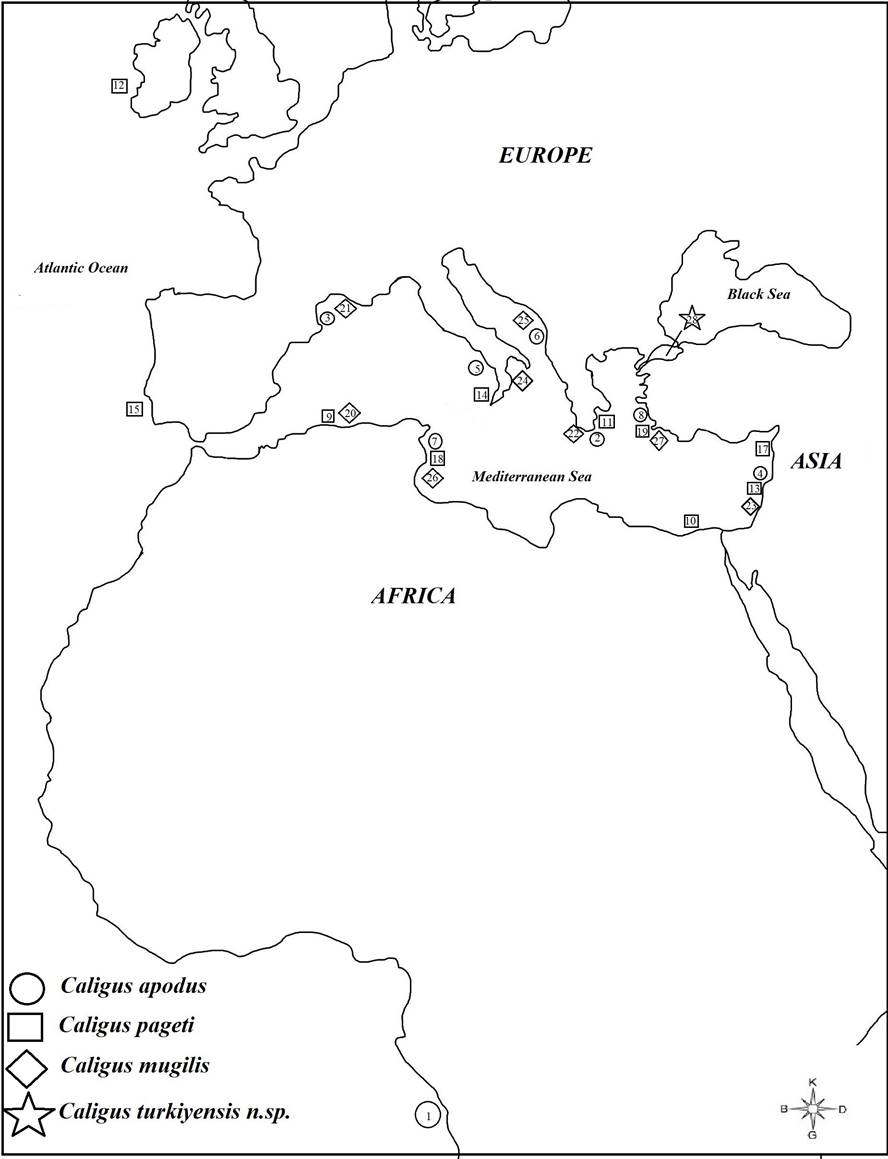
Caligus apodus, C. mugilis and C. pageti have been reported mostly from countries along the Mediterranean coast, except for one report from the Atlantic Ocean (Angola) for C. apodus and from Ireland and South Africa for C. pageti (Fig. 7). Considering the geographical distribution of these parasites, this distribution suggests they are mostly specific to the Mediterranean.
The Sea of Marmara, where the new species discovered in this study was found, is a transitional area between the Black Sea and the Mediterranean, and is under the influence of both these seas. The Sea of Marmara has unique hydrographic, hydrological and oceanographic features. In addition, there is a dense human population around the Sea of Marmara and Bandırma Bay, which are then affected by both domestic and industrial pollution (Öztürk 2021). Due to the unique environmental characteristics of the Sea of Marmara, speciation may have occurred in Caligus parasites in link geographical isolation.
Caligus apodus, C. mugilis and C. pageti have high host specificity and have been reported mostly from 5 species of mugilids: Mugil cephalus, Chelon auratus, C. labrosus, C. ramada (Risso, 1827) and C. saliens (Risso, 1810). In addition, C. apodus has been reported from Crenimugil crenilabis (Forsskål, 1775), Planiliza carinata (Valenciennes, 1836), and Crenimugil seheli (Fabricius, 1775) in Israel by Paperna & Overstreet (1981) (Table III).
Apart from mullets, C. apodus was reported from different hosts, such as the common sole, Solea solea (Linnaeus, 1758) from Iskenderun Bay, Turkey by Özak et al. (2013). The reason why this parasite has been reported from the common sole as well as mullets is probably because the habitats of these hosts are similar. Common sole live in demersal habitats, while C. labrosus and C. saliens live in demersal, M. cephalus in benthopelagic and C. auratus and C. ramada in pelagic-neritic habitats (Froese & Pauly 2024). Apart from pelagic-neritic, the presence of these fish in similar habitats increases the possibility of parasites changing hosts between mullets and common sole.
Venmathi Maran et al. (2016) reviewed the presence and explaining factors of caligid copepods in coastal and marine plankton. Caligids may be found in plankton, because they escape from their host, host switch or search for a new host, change of mode of life, and accidental detachment from the host. This behavior supports the possibility to be found on different hosts.
Due to their host specificity, Boualleg et al. (2011) and Ben Mansour & Ben Hassine (1998) defined C. mugilis and C. pageti as stenoxenous species, respectively. Ragias et al. (2004) only reported it from sea bass, for the same reasons as above.
Caligus minimus has been mainly reported from the North Atlantic Ocean, Mediterranean, Adriatic, Sea of Marmara and Black Sea (Radujkovic & Raibaut 1989, Özak 2020). In addition, there are reports of C. minimus from perciforms other than sea bass, such as Alosidae, Moronidae, Gadidae, Labridae, Mugilidae, Sparidae, Percidae, and Sciaenidae.
The geographical distribution of C. curtus includes the Arctic-Boreal region of the Atlantic Ocean and contiguous waters. It is mainly a typical parasite of marine gadids, but has been reported from marine fish from various families, including elasmobranchs (see Parker et al. 1968). Considering the host specificity of C. curtus and C. minimus, they may be defined as euryxenous parasite species.
The number of studies on species of Caligus in the Mediterranean and Aegean coasts of Turkey is higher than in the Marmara Sea and Black Sea. Twenty of Caligus have been recorded hitherto from Turkish Coasts, of which 19 are from Mediterranean and Aegean Coasts, 5 from Sea of Marmara and 1 from Black Sea Sea (Özak 2020). This parasitological study still includes species of Caligus of mullets in a limited region. The discovery of Caligus turkiyensis n.sp. increases the 21st species of Caligus for Turkish fauna.
Information about the host specificity, host-parasite relationship and zoogeography of parasites of mullets is very important from zoological and ecological point of view. Parasites cause mass and serious fish deaths (Paperna & Lahav 1974). Infections by monoxenous parasites such as monogenea, parasitic crustaceans are more effective in farmed fish (Bragoni et al. 1984, Paperna 1987, Stella et al. 2023). Mullet is the close contact of with farmed fish, which can transmit pathogens such as parasites, viruses, bacteria, etc. to these fish and cause infection, leading to serious economic losses (Johnson et al. 2004, Paladini et al. 2017, Assane et al. 2022). It is also important to identify parasites for fish farming to take precautions against possible infections in the future
CONCLUSION
Although Caligus apodus and C. pageti were known as specific to the coasts of the Mediterranean Sea and Aegean Sea, there are uncertainties about the morphological characters and geographic distribution of the caligids in the Sea of Marmara. This study presented C. apodus, C. pageti and the morphological characters of a new species
Caligus turkiyensis n. sp. from the sea of Marmara. Although morphological characters of this new species are described in this study, molecular studies are needed to confirm its species status.
Acknowledgements: The first author gratefully acknowledges the following individuals for technical and facility support: Dr. Mike Robertson for scientific support, Ömer Öktener who provided the phase contrast condenser for Olympus CH30 Light microscope, Dr. Argun Özak for criticism, fisherman Nilufer Torpilli for helping to catch the fishes, mechanical engineer Mr. Vedat Kızılkaya for repairing the stereomicroscope, Dr. Olivier Brosseau for obtaining an article about Caligus dicentrarchi and Dr. Cigdem Akduman for providing the chemical solutions, the anonymous reviewers and Dr. Yves Desdevises, Dr. Elodie Magnanou, Dr. Franck Lartaud the editors, and Mrs Véronique Arnaud, Editorial Secretary of Vie et Milieu, for the valuable, critical and helpful comments and corrections on the manuscript. This article is dedicated to Prf.Dr. Zbigniew Kabata, Prf.Dr. André Raibaut, and Prf.Dr. Murat Sezgin
| Caligid Species | Host | Locality | References |
| Caligus apodus | Chelon labrosus | France, Italy, Montenegro, Tunisia, Turkey | Raibaut & Ben Hassine (1977), Altunel (1983), Radujkovic & Raibaut (1987), Merella & Garippa (2001), Braun (1981) |
| Chelon auratus | Israel, Tunisia, Turkey | Raibaut & Ben Hassine (1977), Paperna & Overstreet (1981), present study | |
| Chelon ramada | France, Israel, Italy, Tunisia, Turkey | Raibaut & Ben Hassine (1977), Altunel (1983), Merella & Garippa (2001), Paperna & Overstreet (1981), Braun (1981) | |
| Chelon saliens | France, Israel, Italy, Montenegro, Tunisia, Turkey | Raibaut & Ben Hassine (1977), Altunel (1983), Radujkovic & Raibaut (1987), Merella & Garippa (2001), Paperna & Overstreet (1981), Braun (1981) | |
| Mugil cephalus | Angola, France, Israel, Italy, Tunisia, Turkey | Capart (1959), Raibaut & Ben Hassine (1977), Altunel (1983), Merella & Garippa (2001), Paperna & Overstreet (1981), Braun (1981) | |
| Oedalechilus labeo | Montenegro | Radujkovic & Raibaut (1987) | |
| Crenimugil crenilabis | Israel | Paperna & Overstreet (1981) | |
| Planiliza carinata | Israel | Paperna & Overstreet (1981) | |
| Crenimugil seheli | Israel | Paperna & Overstreet (1981) | |
| Dicentrarchus labrax | Greece | Ragias et al. (2004) | |
| Solea solea | Turkey | Özak et al. (2013) | |
| Caligus pageti | Chelon labrosus | Ireland, Israel, Tunisia, Turkey | Paperna (1975), Raibaut & Ben Hassine (1977), Altunel (1983), Holmes (1998) |
| Chelon auratus | Algeria, Israel, Italy, South Africa, Tunisia | Brian (1931) cited by Ramdane & Trilles (2007), Raibaut et al. (1971),Oldewage & van As (1989), Merella & Garippa (2001), Dzikowski et al. (2003) | |
| Chelon saliens | Algeria, Italy, Tunisia, Turkey | Raibaut et al. (1971), Altunel (1983), Merella & Garippa (2001), Boualleg et al. (2010) | |
| Mugil cephalus | Israel, South Africa, Syria, Tunisia, Turkey | Raibaut & Ben Hassine (1977), Paperna & Lahav (1974), Altunel (1983), Hassan et al. (2022), present study | |
| Chelon ramada | Egypt, Israel, South Africa, Tunisia, Turkey | Russell (1925), Paperna (1975), Raibaut & Ben Hassine (1977), Altunel (1983), Oldewage & van As (1989) | |
| Dicentrarchus labrax | Portugal, Greece | Santos (1996), Ragias et al. (2004) | |
| Caligus mugilis | Mugil cephalus | Algeria, Italy | Brian (1906), Boualleg et al. (2010) |
| Chelon labrosus | Algeria, France, Israel, Italy, Montenegro, Tunisia, Turkey | Boualleg et al. (2010), Merella & Garippa (2001), Raibaut & Ben Hassine (1977), Paperna & Overstreet (1981), Ben Hassine (1983), Braun (1981), Radujkovic & Raibaut (1987) | |
| Chelon auratus | Algeria | Boualleg et al. (2010) | |
| Chelon ramada | Algeria | Boualleg et al. (2010) | |
| Dicentrarchus labrax | Greece | Ragias et al. (2004) | |
| Caligus turkiyensis n. sp. | Mugil cephalus | Turkey | present study |
n. sp. | |||||
Female Length of total body | 3.79 mm (3.13-4.61) | 5.33 mm (5.26- 5.4) | 4.38 mm (4.36-4.40) | 8.7 mm (5.1-10.1) | 6.2 mm |
Genital complex length/ Abdomen length | 1.6: 1 | 3.2: 1 | 2.3: 1 | 2.5: 1 | 2.6: 1 |
Number of segment in abdomen | 2 (indistinctly) | 1 | 1 | 1 | 1 |
Length of tines in the sternal furca | Little divergent, slightly elongated | slightly divergent short | Slightly divergent short | Slightly divergent, short parallel | Short parallel |
Number of teeth on mandible | 13 teeth | 12 teeth | 12 teeth | 12 teeth | 12 teeth |
Posterolateral corners of genital complex | Indistinctly lobate, not extend to anterior of the abdomen | Distinctly lobate, extend slightly beyond the middle of abdomen | Distinctly lobate, extend slightly beyond one third of abdomen | Distinctly lobate, extend slightly beyond one third of abdomen | Distinctly lobate, extend slightly beyond one third of abdomen |
Terminal claw of antenna | Slightly recurved | Slightly recurved | Strongly recurved | Strongly recurved | Slightly recurved |
Postantennal process | Slightly recurved | Slightly recurved | Strongly recurved | Strongly recurved | Slightly recurved |
Length/ Anterior margin of maxillule endopodite | 1.45 times | 1.1 times | 1.6 times | 0.8 times | 1.2 times |
| Process on myxal surface of maxilliped | A slight process | Absent | Absent | A slight process | Absent |
| First spine/ Third spine on fourth leg | 3.2 times | 3.4 times | 5 times | 5 times | 4.5 times |
| Denticles on first spine of fourth leg | Absent | Unknown | Absent | Present | Absent |
Male Length of total body | 3.67 mm (3.30-4.69) | Unknown | 4-5mm | 7.93 mm (5.38-12.3) | 6.5 mm |
| Process on myxal surface of maxilliped | One small process in distal and one large bifid process in proximal | Unknown | One large process in proximal | One large process in both proximal and distal | One large process in proximal |
Terminal segment of antenna | Quadrafid plate and a basal seta | Unknown | Quadrafid plate | Bifid plate and a basal seta | Trifid plate |
1
; 2From Raibaut & Ben Hassine (1977)
; 3From Raibaut & Ben Hassine (1977) and Kabata (1979)
From Parker et al. (1968); 4From Cabral & Raibaut (1986)
REFERENCES
Alaş A, Öktener A 2017. Confirmed occurrence of Caligus mugilis Brian (Copepoda, Siphonostomatoida; Caligidae) with morphological characters in Turkish marine waters. Thalassas 33: 133-137. https://doi.org/10.1007/s41208-017-0030-5Altunel FN 1983. Parasitism in Mullets (Mugil spp). I. National Congress of The Marine and Freshwater Researchs. J Ege Univ Sci (B): 364-378.Assane IM, Prada-Mejia KD, Gallani SU, Weiser NF, Valladão GMR, Pilarski F 2022. Enterogyrus spp. (Monogenea: Ancyrocephalinae) and Aeromonas jandaei co-infection associated with high mortality following transport stress in cultured Nile tilapia. Transbound Emerg Dis 69(4): e276-e287. doi: 10.1111/tbed.14295. Epub 2021 Sep 7Ben Hassine OK 1983. Les copépodes parasites de Poissons Mugilidae en Méditerranée occidentale (côtes Francaises et Tunisiennes). Morphologie, Bio-écologie, Cycles évolutifs. Dissertation, Univ. Montpellier II, France, 452 p.Ben Mansour B, Ben Hassine OK 1998. Preliminary analysis of parasitic copepod species richness among coastal fishes of Tunisia. Ital J Zool 65: 341-344. https://doi org/10.1080/11250009809386844Bere R 1936. Parasitic copepods from Gulf of Mexico fish. Am Midl Nat 17(3): 577-625. https://doi.org/10.2307/2419936Boualleg C, Seridi M, Kaouachi N, Quiliquini Y, Bensouilah M 2010. Les Copépodes parasites des Poissons téléostéens du littoral Estalgérien. Bull Inst Sci Rabat Sect Sci de la Vie 32: 65-72.Boualleg C, KaouachiN, Seridi M, Ternengo S, Bensouilah MA 2011. Copepod parasites of gills of 14 teleost fish species caught in the gulf of Annaba (Algeria). Afr J Microbiol Res 5: 425-429. https://doi.org/10.5897/AJMR10.15.Boxshall GA 1990. The skeletomusculature of siphonostomatoid copepods, with an analysis of adaptive radiation in structure of the oral cone. Philos Trans R Soc Lond Ser B Biol Sci 328: 167-212.https://doi.org/10.1098/rstb.1990.0113.Boxshall GA 2018. The sea lice (Copepoda: Caligidae) of Moreton Bay (Queensland, Australia) with descriptions of thirteen new species. Zootaxa 4398: 1-172. https://doi.org/10.11646/zootaxa.4398.1.1Bragoni G, Romestand B, Trilles JP 1984. Parasitoses à cymothoadien chez le Loup, Dicentrarchus labrax (Linnaeus, 1758) en élevage. I. Ecologie parasitaire dans le cas de l’Etang de Diana (Haute Corse) (Isopoda, Cymothoidae). Crustaceana 47: 44-51. https://doi.org/10.1163/156854084X00306Braun M 1981. Contributiono à l’étude biologique des zones à salinité variable du littoral méditerranéen français Copépodes parasites de Mugilidés. These 3ème cycle. Univ Montpellier II, France, 88 p.Brian A 1906. Copepodi parassiti dei Pesci d’Italia. Stab. Tipo-Litografico R. Istituto Sordomuti, Genova, 187 p. https://doi.org/10.5962/bhl.title.58642Brian A 1924. Arthropoda (Première partie) Copepoda. Copépodes commensaux et parasites des côtes mauritaniennes. Parasitologia mauritanica. Bull Com Étud Hist Scient Afr Occid Fr: 365-427.Brian A 1931. Sur la synonymie de Caligus argilasi Brian, 1931 avec le Caligus pageti Russel, 1925. Bull Soc Hist Nat Afrique Nord 22(6): 157.Brian A 1935. I Caligus parassiti dei pesci del Mediterraneo (Copepodi). Ann Mus Civ Stor Nat Giacomo Doria 57(7): 152-211.Cabral P, Raibaut A 1986. Découverte d'un copépode caligide nouveau, parasite du tégument du Loup, Dicentrarchus labrax (L., 1758) (Pisces, Moronidae) en élévage et en milieu naturel. Bull Soc Zool Fr 111(1-2): 123-130.Capart A 1959. Copépodes parasites. Résultats Scientifiques de l'Expédition Océanographique Belge dans les Eaux Côtieres Afrricaines de l'Atlantique Sud (19481949). Inst Royale Sci Nat Belg 3(5): 59-126.Dippenaar SM 2005. Reported siphonostomatoid copepods parasitic on marine fishes of southern Africa. Crustaceana 77(11):1281-1328. https://doi.org/10.1163/1568540043165985Dzikowski R, Paperna I, Diamant A 2003. Use of fish parasite species richness indices in analyzing anthropogenically impacted coastal marine ecosystems. Helgol Mar Res 57(3-4): 220227. https://doi.org/10.1007/s10152-003-0138-2Froese R, Pauly D. 2024. FishBase. World Wide Web electronic publication. www.fishbase.org, version (10/2023).Grobler NJ 2004. A review of the genus Caligus (Copepoda: Caligidae) from South Africa. PhD thesis, University of the Free State, Bloemfontein, South Africa, 203 p.Hamdi I, Benmansour B, Zouari-Tlig S, Kamanli SA, Özak AA, Boxshall GA 2021. Caligus tunisiensis n. sp. (Copepoda: Caligidae) parasitic on the painted comber Serranus scriba (L.) (Perciformes: Serranidae) from the Mediterranean Sea, off the Tunisian coast. Syst Parasitol 98: 5771. https://doi.org/10.1007/s11230-020-09959-9Hassan M, Dayoub A, Gnede S 2022. First record of Caligus pageti (Copepoda: Caligidae) on Mugil cephalus in Syrian marine waters. Tishreen Univ J Res Sci Stud Biol Sci 44(4): 97-103.Hewitt GC 1971. Two species of Caligus (Copepoda, Caligidae) from Australian waters, with a description of some developmental stages. Pac Sci 25(2): 145-164.Ho JS, Lin CL 2003. Solution to the taxonomic confusion surrounding Caligus epinepheli Yamaguti, a caligid copepod (Siphonostomatoida) parasitic on marine fishes. Zool Res 42(2): 268-283.Ho JS, Lin CL 2004. Sea lice of Taiwan (Copepoda: Siphonostomatoida: Caligidae). The Sueichan Press, Keelung, Taiwan. 388 p.Holmes JMC 1998. A checklist of the Siphonostomatoida (Crustacea: Copepoda) of Ireland. Bull Ir Biogeogr Soc 22(2): 194-228.Huys R, Boxshall GA 1991. Copepod Evolution. London: The Ray Society.Johnson SC, Treasurer JW, Bravo S, Nagasawa K, Kabata Z 2004. A review of the impact of parasitic copepods on marine aquaculture. In: Hwang JS, JS Ho, CT Shih, (Eds). Proceedings of the 8th International Conference on Copepoda, Special Issue: Contemporary Studies on Copepoda. Zoological Studies 43(2): 229-243.Kabata Z 1965. Copepoda parasitic on Australian fishes. IV. Genus Caligus (Caligidae). Ann Mag Nat Hist 13(8): 109-126. https://doi.org/10.1080/00222936508651546Kabata Z 1979. Parasitic Copepoda of British Fishes, Ray Society Publications, The Bristish Museum, London, 152, 468 p.Kabata Z 2003. Copepods Parasitic on Fishes. 2nd, revised edition. Synopses of the British Fauna, 2nd revised ed. Backhuys. No 47: 1-274.Merella P, Garippa G 2001. Metazoan parasites of grey mullets (Teleostea: Mugilidae) from the Mistras Lagoon (Sardinia, western Mediterranean). Sci Mar 65(3): 201-206. https://doi.org/10.3989/scimar.2001.65n3201Müller OF 1785. Entomostraca seu Insecta Testacea, quae in aquis Daniae et Norvegiae reperit, descripsit et iconibus illustravit Otho Fridericus Müller. F.W. Thiele, Lipsiae & Havniae: 1-134.Ohtsuka S, Boxshall GA 2019. Two new species of the genus Caligus (Crustacea, Copepoda, Siphonostomatoida) from the Sea of Japan, with a note on the establishment of a new species group. ZooKeys 893: 91-113. https://doi.org/10.3897/zookeys.893.46923Ohtsuka S, Nawata M, Nishida Y, Nitta M, Hirano K, Adachi K, Kondo Y, Venmathi Maran BA, Suárez Morales E 2020. Discovery of the fish host of the ‘planktonic’ caligid Caligus undulatus Shen & Li, 1959 (Crustacea: Copepoda: Siphonostomatoida). Biodivers Data J 8: e52271. https://doi.org/10.3897/BDJ.8.e52271Oldewage WH, van As JG 1989. Occurrence and distribution of Caligus (Copepoda: Siphonostomatoida) in African coastal waters. Rev Zool Afric 103(1): 91-98.Otto AG 1821. Conspectus animalium quorundam maritimorum nondum editorum pars prior quam patri dilectissimo ad cineres usque venerando Bernhardo Crristiano Otto pro celebrando die laetissimo XI. Maji MDCCCXXI quo ante decem lustra honores doctorales acceperat. Typis Universitatis, Vratislaviae, 20 p.Öktener A, Türker D, Alaş A 2017. The Sea of Marmara: New Locality for Two Caligids and one Lernanthropid in Turkey. J Wetl Biodivers 7: 109-30.Özak AA 2020. Sea lice (Copepoda: Caligidae) of Turkey, with the discovery of Caligus quadratus Shiino, 1954 in the Mediterranean Sea and the re-description of a rare caligid copepod, Caligus scribae Essafi, Cabral & Raibaut, 1984. Syst Parasitol 97(6): 779-808. https://doi.org/10.1007/s11230-020-09953-1.Özak AA, Demirkale İ, Boxshall GA, Etyemez M 2013. Parasitic copepods of the common sole, Solea solea (L.), from the Eastern Mediterranean coast of Turkey. Syst Parasitol 86: 173-185. https://doi.org/10.1007/s11230-013-9441-8.Öztürk İ 2021. The Marmara Sea & The Turkish Straits System in Light of Scientific Data. Ankara: TurkishAcademyofSciences, vol31: 190p.Paladini G, Longshaw M, Gustinelli A, Shinn AP. 2017. Parasitic diseases in aquaculture: their biology, diagnosis and control. In: Austin B, Newaj-Fyzul A, editors. Diagnosis and control of diseases of fish and shellfish. Hoboken: Chichester (UK): John Wiley & Sons Ltd. p. 37-107.https://doi.org/10.1002/9781119152125.ch4Paperna I 1975. Parasites and diseases of the grey mullet (Mugilidae) with special reference to the seas of the Near East. Aquaculture 5(1): 6580. https://doi.org/10.1016/0044-8486(75)90018-6.Paperna I 1987. Solving parasite-related problems in cultured marine fish. Int J Parasitol 17: 327-336.https://doi.org/10.1016/0020-7519(87)90107-XPaperna I, Lahav M 1974. Mortality among grey mullets in a seawater pond due to caligid parasitic copepod epizootic. Bamidgeh, Bull Fish Cult Israel 26(1): 12-15.Paperna I, Overstreet R 1981. Parasites and Diseases of Mullets (Mugilidae). Fac Public Harold W. Manter Labo Parasitol 579: 411-493.Parker RR, Kabata Z, Margolis L, Dean MD 1968. A review and description of Caligus curtus Müller, 1785 (Caligidae: Copepoda), type species of its genus. J Fish Res Board Can 25(9): 1923-1969. https://doi.org/10.1139/f68-172Pritchard MH, Kruse GO 1982. The collection and preservation of animal parasites. Technical Bulletin No. 1. The Harold W. Manter Laboratory, University of Nebraska Press, 141 p.Radujkovic BM, Raibaut A 1987. Copépodes parasites des Poissons des côtes du Monténégro (Adriatique sud). 1er série. Acta Adriat 28(1-2): 121-142.Radujkovic BM, Raibaut A 1989. Parasites des Poissons marins du Monténégro: Copépodes. Acta Adriat 30(1-2): 237-278.Ragias V, Tontis D, Athanassopoulou F 2004. Incidence of an intense Caligus minimus Otto 1821, C. pageti Russel, 1925, C. mugilis Brian, 1935 and C. apodus Brian, 1924 infection in lagoon cultured sea bass (Dicentrarchus labrax L.) in Greece. Aquaculture 242(1-4): 727-733. https://doi.org/10.1016/j.aquaculture.2004.08.019Raibaut A, Ben Hassine OK, Maamouri K 1971. Copépodes parasites des Poissons de Tunisie (première série). Bull Inst Natl Sci Tech Oceanogr Peche Salammbo 2(2): 169-197.Raibaut A, Ben Hassine OK 1977. Les copépodes parasites des Muges en Méditerranée. Bull Mus Natl Hist Nat 472 (Zool. 329): 833-848.Raibaut A, Combes C, Benoit F 1998. Analysis of the parasitic copepod species richness among Mediterranean fish. J Mar Syst 15: 185-206. https://doi.org/10.1016/S0924-7963(97)00079-1Ramdane Z, Trilles JP 2007. Parasitic copepods (Crustacea: Copepoda) from Algerian marine fishes. Zootaxa 1574: 49-68. https://doi. org/10.11646/zootaxa.1574.1.3Russel FS 1925. A new species of Caligus from Egypt, Caligus pageti, sp. n. Ann Mag Nat Ilist 15(9): 611-618. https://doi.org/10.1080/00222932508633257Santos MJ 1996. Observations on the Parasitofauna of wild seabass (Dicentrarchus labrax L.) from Portugal. Bull Eur Assoc Fish Pathol 16(3): 77-79.Skinner RH 1975. Parasites of the striped mullet, Mugil cephalus, from Biscayne Bay, Florida, with descriptions of a new genus and three new species of Trematodes. Bull Mar Sci 25(3): 318-345.Soler-Jiménez LC, Morales-Serna FN, Aguirre-Macedo ML, McLaughlin JP, Jaramillo AG, Shaw JC, James AK, Hechinger RF, Kuris AM, Lafferty KD, Vidal-Martínez VM 2019. Parasitic copepods (Crustacea, Hexanauplia) on fishes from the lagoon flats of Palmyra Atoll, Central Pacific. ZooKeys 833: 85-106. https://doi.org/10.3897/zookeys.833.30835Stella E, Pastres R, Pasetto D, Kolega M, Mejdandžić D, Čolak S, Musmanno A, Gustinelli A, Mari L, Bertuzzo E 2023. A stratified compartmental model for the transmission of Sparicotyle chrysophrii (Platyhelminthes: Monogenea) in gilthead seabream (Sparus aurata) fish farms. R Soc Open Sci 10: 221377. https://doi.org/10.1098/rsos.221377Urawa S, Kato T 1991. Heavy infections of Caligus orientalis Gussev (Copepoda: Caligidae) on caged rainbow trout Oncorhynchus mykiss in brackish water. Gyobyo Kenkyu (Fish Pathol) 26: 161-162.Venmathi Maran BA, Suárez-Morales E, Ohtsuka S, Soh HY, Hwang UW 2016. On the occurrence of caligids (Copepoda: Siphonostomatoida) in the marine plankton: a review and checklist. Zootaxa 4174(1): 437-447. https://doi.org/10.11646/zootaxa.4174.1.27Walter TC, Boxshall G 2024. World of Copepods Database. Caligus Müller O.F., 1785. Accessed through: World Register of Marine Species at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=135566 on 2024-02-12Wilson B 1908. North American parasitic copepods: new genera and species of Caliginae. through: World Register of Marine Species at:https://www.marinespecies.org/aphia.php?p=taxdetails&id=135566 on 2024-02-12Wilson B 1908. North American parasitic copepods: new genera and species of Caliginae. Proc US Natl Mus 33(1580): 593-627. https://doi.org/10.5479/si.00963801.33-1580.593WoRMS 2024. World Register of Marine Species. Available from https://Accessed 2024-02-12. doi:10.14284/170Received on July 29, 2023
Accepted on April 4, 2024
Associate editor: Y. Desdevises